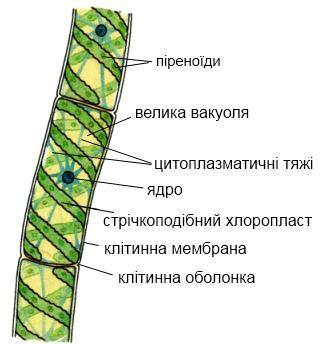
Будова спірогири. (Рис.1)
Талом водорості виглядає як нитка, що не гілкується та складається з одного ряду однакових клітин циліндричної форми. Клітини відносно великі, довжина їх може досягати в деяких видів до 0,01 мм. Ширина нитки спірогири становить від 5 до 200 мкм.
Оболонкою кожної клітини є целюлозна клітинна стінка, покрита зовні слизом. Більшу частину клітини займає вакуоль з клітинним соком. На цитоплазматичних тяжах, що проходять через вакуоль, підвішене одне ядро. У кожній клітині є по одному і більше спірально закручених хлоропластів, що нагадують стрічки, розташовані у пристінковому шарі цитоплазми. Всі спірогири - автотрофи, які синтезують органічні речовини на світлі в хлоропластах в процесі фотосинтезу.
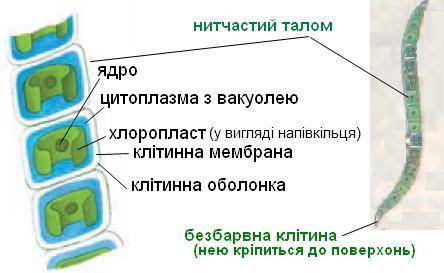
Будова улотрикса. (Рис.2)
Баговиння улотрикса складається з нерозгалужених ниток різної довжини, які на початку росту прикріплюються до субстрату безбарвною видовженою клітиною — ризоїдом. Клітини ниток циліндричні або бочкоподібні, короткі. Кожна клітина має ядро, пристінний хлоропласт у вигляді неповного кільця і один або кілька піреноїдів.
Більшу частину життя водорість проводить у стані гаметоспорофіта, який має вигляд довгих нерозгалужених однорядних ниток. У кожної клітини нитки (за винятком базальної) є пристінний хлоропласт у вигляді незамкненого кільця, кілька піреноїдів, одне ядро; центр клітини зайнятий вакуолею з клітинним соком. Всі клітини здатнi до поділу, що обумовлює постійне наростання талому в довжину, а також утворення репродуктивних клітин. Базальна клітина, за до якої водорiсть прикріплюється до субстрату, відрізняється від звичайних вегетативних клітин нитки: вона витягнута у короткий ризоїд, не здатна до подiлу, має редукований хлоропласт.
Чередование поколений
Чередование поколений, закономерная смена у организмов генераций, различающихся типом размножения.
У животных имеются первичное и вторичное Ч. п. Первичным Ч. п., свойственным многим простейшим, считают смену половой генерации поколением, размножающимся неполовыми клетками (агаметами). Так, у фораминифер (рис. 1) чередующиеся поколения представлены половыми и бесполыми особями — гамонтами и агамонтами. Гамонты в результате многократного деления ядра образуют гаметы, которые, копулируя попарно, дают зиготу, вырастающую в агамонта. Последний в результате шизогонии распадается на агаметы — будущие гамонты. Редукционное деление (мейоз) происходит перед образованием агамет, поэтому половое поколение гаплоидно, так же как и гаметы, тогда как зигота и агамонты диплоидны. У споровиков и жгутиконосцев диплоидна только зигота, поскольку мейоз осуществляется при её первом делении. У солнечников, некоторых жгутиконосцев и инфузорий мейоз связан с образованием гамет, которые являются единственной гаплоидной стадией жизненного цикла. Такие же отношения свойственны всем многоклеточным животным. Вторичное Ч. п. встречается у животных в двух формах. Чередование различных форм полового размножения, например нормального полового процесса с партеногенезом, называется гетерогонией, а чередование полового размножения с бесполым посредством многоклеточных вегетативных тел или путём поперечного деления — метагенезом. Гетерогония характерна для трематод, некоторых круглых червей и коловраток, а также для ряда членистоногих — дафний, тлей, орехотворок, некоторых мух-галлиц и др. Метагенез очень характерен для оболочников (сальп, бочёночников, асцидий, пиросом) и кишечнополостных (гидроидных и сцифоидных), у которых половое поколение представлено одиночными свободноплавающими медузами, а бесполое — сидячими полипами, нередко образующими