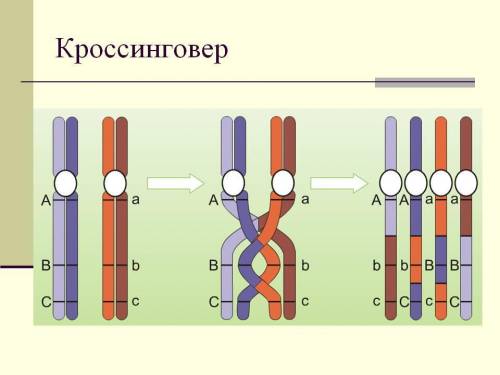
1. Что является причиной нарушения сцепления генов?
Причиной нарушения сцепления генов является кроссинговер (обмен участками гомологичных хромосом), который происходит в профазе мейоза 1
2. Каково биологическое значение обмена аллельными генами между гомологичными хромосомами?
Обмен аллельными генами приводит к появлению новых сочетаний генов, что дает материал для генетической изменчивости. Возникают новые сочетания признаков
3. Подтверждена ли цитологически теория сцепленного наследования?
Подтверждена. В ходе клеточного деления хромосомы в анафазе расходятся к полюсам клетки целиком, а следовательно гены должны наследоваться группой - сцепленно. Однако иногда сцепленность нарушается, так как в ходе профазы 1 мейоза происходит кроссинговер - обмен участками гомологичных хромосом
Синтетическая теория в её нынешнем виде образовалась в результате переосмысления ряда положений классического дарвинизма с позиций генетики начала XX века. После переоткрытия законов Менделя (в 1901 г.), доказательства дискретной природы наследственности и особенно после создания теоретической популяционной генетики трудами Рональда Фишера, Джона Б. С. Холдейна-младшего и Сьюэла Райта, учение Дарвина приобрело прочный генетический фундамент.
Влияние генов на строение и функции организма плейотропно: каждый ген участвует в определении нескольких признаков. С другой стороны, каждый признак зависит от многих генов; генетики называют это явление генетическойполимерией признаков. Фишер говорит о том, что плейотропия и полимерия отражают взаимодействие генов, благодаря которому внешнее проявление каждого гена зависит от его генетического окружения. Поэтому рекомбинация, порождая всё новые генные сочетания, в конце концов создает для данной мутации такое генное окружение, которое позволяет мутации проявиться в фенотипе особи-носителя. Так мутация попадает под действие естественного отбора, отбор уничтожает сочетания генов, затрудняющие жизнь и размножение организмов в данной среде, и сохраняет нейтральные и выгодные сочетания, которые подвергаются дальнейшему размножению, рекомбинации и тестированию отбором. Причем отбираются прежде всего такие генные комбинации, которые благоприятному и одновременно устойчивому фенотипическому выражению изначально мало заметных мутаций, за счет чего эти мутантные гены постепенно становятся доминантными. Эта идея нашла выражение в труде Р. Фишера «The genetical theory of natural selection» (1930). Таким образом, сущность синтетической теории составляет преимущественное размножение определённых генотипов и передача их потомкам. В вопросе об источнике генетического разнообразия синтетическая теория признает главную роль за рекомбинацией генов.
Считают, что эволюционный акт состоялся, когда отбор сохранил генное сочетание, нетипичное для предшествующей истории вида. В итоге для осуществления эволюции необходимо наличие трёх процессов:
мутационного, генерирующего новые варианты генов с малым фенотипическим выражением; рекомбинационного, создающего новые фенотипы особей; селекционного, определяющего соответствие этих фенотипов данным условиям обитания или произрастания. Ф. Добржанского, Дж. Хаксли, Э. Майра, Б. Ренша, Дж. Стеббинса. Это, конечно, далеко не полный список. Только из русских учёных, по меньшей мере, следовало бы назвать И. И. Шмальгаузена, Н. В. Тимофеева-Ресовского, Г. Ф. Гаузе, Н. П. Дубинина, А. Л. Тахтаджяна. Из британских ученых велика роль Дж. Б. С. Холдейна-младшего, Д. Лэка, К. Уоддингтона, Г. де-Бира. Немецкие историки среди активных создателей СТЭ называют имена Э. Баура, В. Циммермана, В. Людвига, Г. Хеберера и других.
У голосеменных растений пыльца формируется в пыльцевых мешках мужских шишек Мужские гаметы (два спермия) образуются при делении генеративной клетки. Из вегетативной клетки образуется пыльцевая трубка, по которой продвигаются спермии. Женский гаметофит у голосеменных растений образуется в семязачатках, которые находятся на чешуях женских шишек. Из четырех образовавшихся в результате мейоза мегаспор одна прорастает (три остальные редуцируются) и образует ткань женского эндосперма (первичного эндосперма) . Это и есть женский гаметофит голосеменных растений. В ткани эндосперма образуются сильно редуцированные женские органы – архегонии, в которых и развиваются женские гаметы – яйцеклетки У покрытосеменных семязачатки (или семяпочки) находятся внутри завязи. В них также образуются четыре мегаспоры, из которых три редуцируются, а оставшаяся делится несколько раз митозом и образует восьмиядерный зародышевый мешок – женский гаметофит покрытосеменных растений. В зародышевом мешке одна из клеток превращается в женскую гамету – яйцеклетку, два гаплоидных ядра сливаются в центре зародышевого мешка и образуют диплоидную центральную клетку, а остальные клетки исчезают
Объяснение: